Dracoraptor hanigani (terópodo) restaurado como depredador y carroñero que habita en la costa. Ilustraciones de Bob Nicholls (paleocreations.com). Crédito:Museo de Historia Natural, Londres. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
La evolución de la endotermia (termorregulación por medios metabólicos) representa una transición importante en la historia de los vertebrados. Sin embargo, el proceso de evolución de la endotermia y su cronología en aves y mamíferos sigue siendo controvertido. En un nuevo informe sobre Avances de la ciencia , Enrico L. Rezende y un equipo de investigadores del Centro de Ecología Aplicada y Sostenibilidad, y el Instituto de Ciencias Ambientales y Evolutivas de Chile, combinó un modelo de transferencia de calor con datos de tamaño corporal de terópodos. Luego, los investigadores reconstruyeron la evolución de las tasas metabólicas a lo largo del linaje del tallo de las aves. Los resultados sugirieron que una reducción en el tamaño constituía el camino de menor resistencia para que evolucionara la endotermia, maximizando la expansión del nicho térmico, mientras se reducen los costos de los elevados requisitos de energía.
Como resultado, los investigadores plantean la hipótesis de que el metabolismo habría aumentado con la miniaturización durante el período Jurásico Temprano-Medio (hace aproximadamente 180 a 170 millones de años) para dar como resultado un gradiente de niveles metabólicos en la filogenia de terópodos. Si bien los terópodos basales pueden haber exhibido tasas de metabolismo más bajas, los linajes no aviarios más recientes eran probablemente termorreguladores decentes con un metabolismo mejorado. El análisis proporcionó una secuencia tentativa en el tiempo de las transiciones evolutivas clave, para la aparición de pequeños, dinosaurios endotérmicos y con plumas voladoras.
La evolución de la endotermia en aves y mamíferos es una transición importante durante la evolución de los vertebrados proporcionando un ejemplo extraordinario de convergencia evolutiva entre grupos. fundamental para su amplia distribución geográfica y éxito ecológico. Aunque varios grupos de invertebrados y vertebrados pueden elevar sus temperaturas por encima de la ambiente, la capacidad de mantener una temperatura corporal alta y constante mediante la producción de calor endógeno en reposo es exclusiva de las aves y los mamíferos. La observación explicó su mayor movilidad, resistencia y tolerancia en una variedad de condiciones. Sin embargo, esta estrategia es energéticamente costosa y no tiene rastro virtual en el registro fósil, por lo tanto, el ritmo y el modo de la endotermia sigue siendo controvertido durante la evolución de los vertebrados.
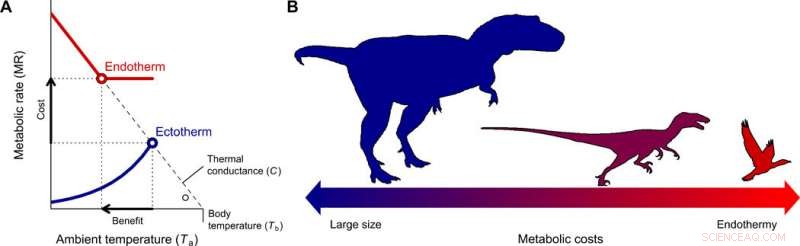
La evolución de la endotermia y la miniaturización en el linaje de terópodos que conducen a las aves. (A) El costo-beneficio de cambiar de ectotermia a endotermia para diferentes rangos de tamaño corporal se cuantificó con el modelo de Scholander-Irving, que describe cómo un aumento en el metabolismo en reposo (costo) aumenta el nicho térmico Tb - Ta (beneficio). Debido a que no existe un gradiente térmico entre el organismo y el medio ambiente en ausencia de producción de calor, esta curva se cruza con la abscisa en Tb =Ta cuando MR =0 (8). Las líneas sólidas azul y roja representan las curvas metabólicas de un ectotermo y endotermo típicos, respectivamente, y los símbolos abiertos representan el gradiente térmico máximo Tb - Ta posible con tasas metabólicas en reposo, utilizado en nuestro modelo (Ec. 2). (B) Una reducción en el tamaño del cuerpo, consistente con el descrito desde terópodos ancestrales hasta aves basales (22), constituye el camino evolutivo de menor resistencia ya que los costos energéticos de ser grande se intercambian por los de ser endotérmico. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
Para comprender el origen y la razón de la endotermia durante la evolución de las aves y los mamíferos, los científicos consideraron dos cuestiones fundamentales. ¿Cuáles son los costos y beneficios de esta estrategia en comparación con la ectotermia? ¿Y qué condiciones favorecieron una transición hacia la endotermia? Rezende y col. abordó las preguntas utilizando el modelo de transferencia de calor de Scholander-Irving, utilizado para estudiar la termorregulación en endotermos durante más de 60 años. La relación rara vez se usó para ectotermos, pero como todos los organismos vivos producen calor endógeno, el modelo siguió siendo aplicable en régimen térmico estable. Hicieron esta suposición crucial para evitar el uso de modelos dinámicos complejos que a menudo se aplican a la ectotermia, lo que podría dificultar los análisis actuales.
Cuantificaron los costos de la endotermia como gasto de energía independiente de la masa, donde los beneficios incluyeron una mayor movilidad y eficiencia de alimentación, evitación de depredadores, tolerancia y colonización de una amplia gama de condiciones ambientales, con mayores tasas de crecimiento y homeostasis. Rezende y col. cuantificó el nicho térmico que podrían ocupar los organismos y su expansión para estimar el beneficio neto de la endotermia. Luego calcularon el costo-beneficio de adoptar un estilo de vida endotérmico con referencia al ancestro ectotérmico y al descendiente endotérmico. Como lo propuso originalmente el biólogo evolutivo Bran K. McNab, cuando los científicos replicaron estos cálculos con estimaciones exactas del tamaño corporal, Los resultados mostraron que los tamaños más pequeños redujeron los costos de energía para evolucionar hacia la endotermia.
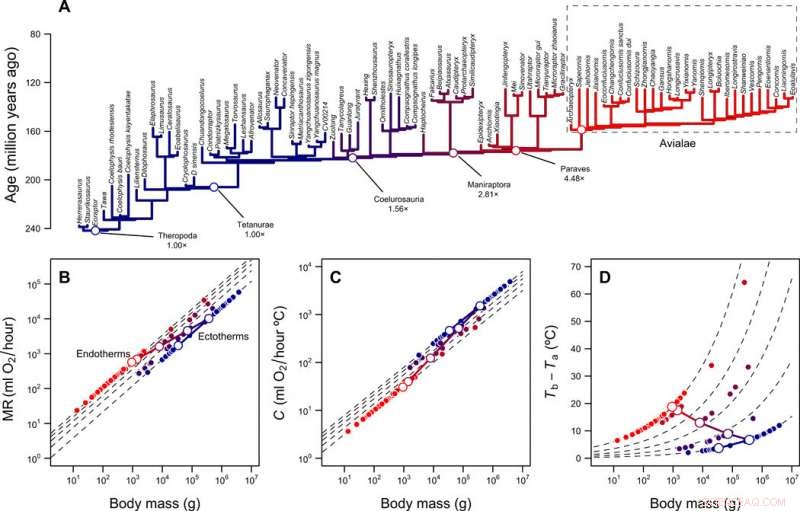
Reconstrucción de niveles metabólicos y nicho térmico de terópodos. (A) Filogenia de terópodos con ramas codificadas por colores según los niveles metabólicos reconstruidos. (B) Escala de la tasa metabólica frente a la masa corporal para ectotermos (MR =0.68mass0.75) y endotermos (MR =3.4mass0.75) y la trayectoria predicha del linaje del tallo de las aves durante la transición de la ectotermia a la endotermia. Las líneas discontinuas muestran diferencias de pliegue entre ectotermos y endotermos (1 × a 5 ×); los símbolos abiertos y cerrados representan valores reconstruidos para el linaje del tallo de las aves y las puntas de la filogenia, respectivamente. (C) Escala de conductancia térmica C y masa corporal para ectotermos (C =2.5mass0.5) y endotermos (C =1.0mass0.5), pliegue las diferencias de 2,5 × a 1 ×. (D) Gradiente térmico y diferencias de pliegue calculadas con la Ec. 1 (en la publicación) y valores en (B) y (C). Las trayectorias lineales log-log que conectan MR y C del ancestro ectotérmico y el descendiente endotérmico, así como la trayectoria resultante en gradiente térmico, se muestran con líneas continuas. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
El equipo exploró cómo este modelo de transferencia de calor se combinó con filogenias y reconstrucciones del tamaño corporal, arrojar luz sobre la evolución de la endotermia en las aves y sus antepasados terópodos. Rezende y col. estimó los costos de la evolución de la endotermia a lo largo del linaje del tallo de las aves utilizando tamaños corporales ancestrales reconstruidos, basado en el registro fósil. Para cuantificar los costos de energía dentro de escenarios alternativos, simularon la evolución del tamaño corporal a lo largo del linaje y obtuvieron la distribución del costo por título en el modelo. Asumieron un modelo de Ornstein-Uhlenbeck (OU) no dirigido entre una masa de 10 gy 100, 000 kg con una tasa de evolución media equivalente a las reportadas para los terópodos. Las simulaciones indicaron una marcada disminución en los costos de energía por grado con la miniaturización. Los científicos explicaron la reducción de costos utilizando dos fenómenos.
Primero, la expansión del nicho térmico basada en un aumento de la tasa metabólica (IM) fue desproporcionadamente mayor en los ectotermos más grandes debido a su capacidad para mantener una temperatura corporal alta (T B ). Esto se acompañó de una tasa metabólica independiente de masa relativamente baja, debido a la homeotermia inicial (termorregulación estable). Respectivamente, notaron que mayor era el tamaño inicial del ancestro ectotérmico, más barata la transición a la endotermia. Segundo, durante la miniaturización, los animales intercambiaron los costos energéticos de ser grandes por ser endotérmicos. Los resultados explicaron cómo evolucionaron las altas tasas de rotación de energía, independientemente de su impacto en las necesidades de alimentos y agua. A pesar de la variación inherente en la disponibilidad de recursos en el tiempo evolutivo, en este linaje se favorecieron sistemáticamente tamaños más pequeños y tasas de rotación de energía más altas. Los resultados del trabajo coincidieron cualitativamente con otros modelos sobre la evolución de la endotermia.
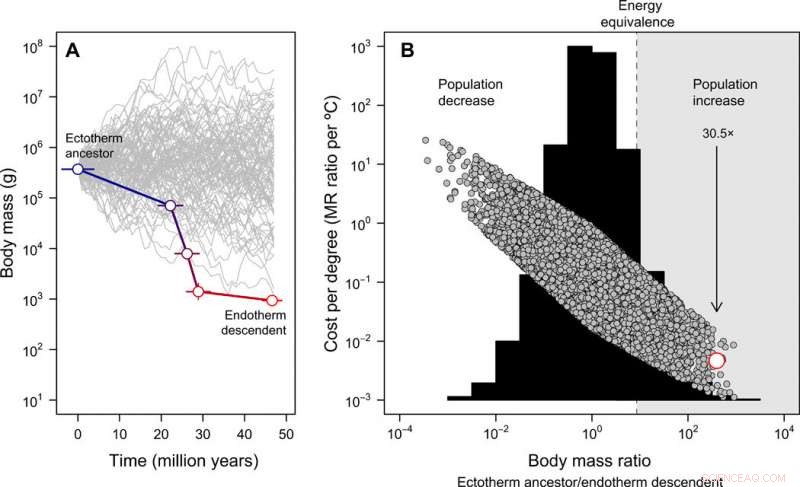
Evolución del tamaño corporal y costo-beneficio de la endotermia. (A) La miniaturización de Tetanurae a aves basales inferida del registro fósil, contrastado con 100 trayectorias de tamaño simulado a partir del mismo tamaño de cuerpo ancestral con fines ilustrativos (tenga en cuenta que para el modelo nulo completo posterior, el tamaño del cuerpo ancestral puede variar). El error representa la DE en valores reconstruidos en 20 árboles candidatos. (B) La distribución de frecuencia de las proporciones de masa corporal obtenidas a través de 10, 000 trayectorias simuladas de tamaño corporal (histograma) y los costos de energía para desarrollar la endotermia expresados por grado Celsius (Ec. 2 en la publicación) bajo este modelo nulo (símbolos grises). En este caso, el tamaño del cuerpo ancestral se obtuvo a partir de una distribución uniforme que oscila entre 10 gy 100, 000 kg. La estimación empírica en el linaje del tallo de las aves se muestra en rojo. La región en la que una reducción en el tamaño corporal compensaría los costos energéticos de la endotermia en evolución, Permitir que la población aumente en un escenario de recursos constantes, se resalta en gris. La flecha muestra el aumento esperado de la población, dada la reducción observada del tamaño corporal en el linaje del tallo de las aves a medida que evolucionaba la endotermia. Estos análisis indican que los costos de energía para desarrollar la endotermia se reducen con la miniaturización y, como resultado, el tamaño de la población puede haber aumentado a pesar de los costos metabólicos de un estilo de vida endotérmico. Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
La reducción de tamaño en el linaje del tallo de las aves, coincidió estrechamente con el camino teórico de menor resistencia para que la endotermia evolucionara, luego Rezende et al. reconstruyó cómo podría haberse desarrollado el fenómeno en la filogenia de los terápodos. El proceso indicó el aumento de MR (tasa metabólica) que abarca la mayor parte del Jurásico Temprano-Medio (hace aproximadamente 180 a 170 millones de años), involucrando grupos de terápodos donde la ocurrencia de protoplumas y plumas ya era omnipresente. También sugirieron tasas metabólicas muy diversas en los linajes concurrentes de Coelurosauria, Maniraptora y Paraves, lo que podría explicar el surgimiento y diversificación de estos grupos durante el período Jurásico tardío. El trabajo también podría explicar la diversidad anormalmente alta de Coelurosauria en tamaños corporales intermedios (30 a 300 kg) en comparación con otros grupos de dinosaurios. El equipo de investigación demostró además un modelo completo de radiación adaptativa en dinosaurios mesozoicos con tasas excepcionales de reducción del tamaño corporal en el linaje del tallo de las aves (es decir, dentro de los nodos basales de Coelurosauria y Paraves), sin embargo, no propusieron una hipótesis evolutiva adecuada para explicar el resultado.
De este modo, Enrico L. Rezende y sus colegas observaron dos fenómenos excepcionales durante la evolución de las aves; (1) una miniaturización sostenida (pero no necesariamente gradual) que abarca millones de años y (2) la aparición de la endotermia. Utilizaron reconstrucciones para sugerir la evolución concomitante de la endotermia con una disminución en el tamaño a lo largo del linaje del tallo de las aves. como también se propuso previamente para los mamíferos. Los correspondientes clados de terápodos exhibieron un espectro completo de MR. Las interpretaciones indican que la endotermia precedió a la evolución del vuelo y la marcada reducción del tamaño corporal durante la evolución del linaje del tallo de las aves estuvo acompañada de un cambio importante en los niveles metabólicos.
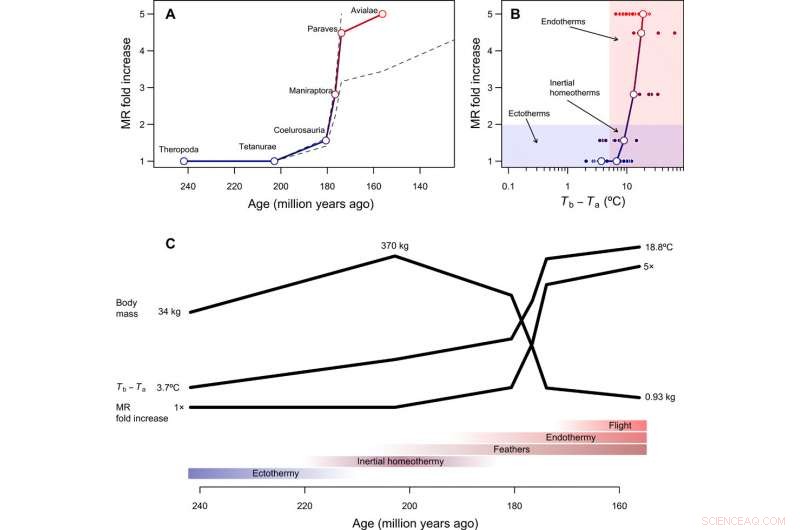
Tempo y modo en la evolución de la endotermia. (A) Curso temporal reconstruido de la evolución metabólica en el linaje del tallo de las aves, con líneas discontinuas que muestran cómo cambian las reconstrucciones suponiendo que Paraves o Neornithes fueran completamente endotérmicos en lugar del pájaro basal [para cálculos con Neornithes, asumimos un tamaño corporal de 150 g basado en estimaciones para Vegavis y una estimación de tiempo de hace 100 Ma]. El aumento de veces en la MR se calculó dividiendo la MR reconstruida durante la transición a la endotermia por la MR esperada para un ectotermo de tamaño similar y, por lo tanto, es adimensional e independiente del tamaño corporal. (B) El camino evolutivo de menor resistencia de la ectotermia a la endotermia incluye la homeotermia inercial como etapa de transición, seguido de un aumento en el metabolismo concomitantemente con una reducción de tamaño. (C) Secuencia hipotética de transiciones evolutivas en el linaje del tallo de las aves, que combina los resultados de este estudio con reconstrucciones filogenéticas de estructuras epidérmicas y capacidad de vuelo activo (ver el texto principal). Crédito:Science Advances, doi:10.1126 / sciadv.aaw4486
El escenario propuesto explicaba por qué los terópodos favorecían sistemáticamente un estilo de vida caro (mayor tasa de rotación de energía) a pesar de sus costos energéticos. y miniaturización sostenida que precede al origen de las aves. Es más, un estado de homeotermia inercial constituía un estado de transición necesario con bajos costos metabólicos. Si los grandes terópodos ancestrales estuvieran fisiológicamente comprometidos con la homeotermia, Rezende y col. luego espere que se hayan apartado de la alometría metabólica ectotérmica (relación entre el tamaño del cuerpo y la forma, anatomía, fisiología y comportamiento) con miniaturización. Durante el tiempo evolutivo, es razonable esperar que los linajes exploten nichos recién abiertos y eventualmente se diversifiquen. Los resultados de este trabajo son preliminares; simplemente destacan que es probable que haya más en la evolución de la endotermia en los arcosaurios, dinosaurios y aves actuales. Los resultados forman una hipótesis de trabajo que se puede combinar en estudios futuros con análisis previos sobre la evolución del tamaño y otras características en el linaje del tallo de las aves para formar interpretaciones bien definidas de secuencias temporales durante las transiciones evolutivas clave.
© 2020 Science X Network