De acuerdo con las leyes básicas de la física, todos los seres vivos necesitan energía del medio ambiente de alguna forma para mantener la vida. Claramente, diferentes organismos han desarrollado diversos medios para recolectar combustible de diversas fuentes para impulsar la maquinaria celular que impulsa procesos cotidianos como el crecimiento, la reparación y la reproducción.
Las plantas y los animales obviamente no adquieren alimentos (o su equivalente en organismos que en realidad no pueden "comer" nada) por medios similares, y sus respectivas entrañas no digieren las moléculas extraídas de fuentes de combustible de forma remota de la misma manera. Algunos organismos requieren oxígeno para sobrevivir, otros son asesinados por él y otros pueden tolerarlo pero funcionan bien en su ausencia.
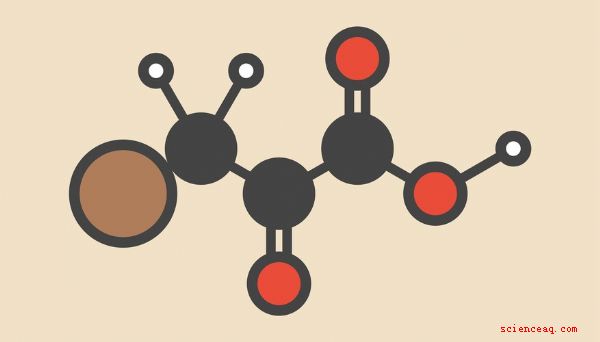
A pesar de la variedad de estrategias que los seres vivos emplean para extraer energía de los enlaces químicos en el carbono. compuestos ricos, la serie de diez reacciones metabólicas denominadas colectivamente glucólisis Una descripción general de las principales entradas y salidas de la glucólisis es un buen punto de partida para comprender cómo las células convierten las moléculas reunidas del mundo externo en energía para sostener la miríada de procesos vitales. en el que las células de su cuerpo están continuamente involucradas. Los reactivos de la glucólisis a menudo se incluyen en la glucosa y el oxígeno, mientras que el agua, el dióxido de carbono y el ATP (trifosfato de adenosina, la molécula que vive más comúnmente para alimentar los procesos celulares) se administran como productos de glucólisis, como sigue: C 6H 12O 6 + 6 O 2 -> 6 CO 2 + 6 H 2O + 36 (o 38) ATP Llamar a esto "glucólisis", como hacen algunos textos, es incorrecto. Esta es la reacción neta de la respiración aeróbica en su conjunto, de la cual la glucólisis es el paso inicial. Como verá en detalle, los productos de la glucólisis per se son en realidad piruvato y una cantidad modesta de energía en forma de ATP: C 6H 12O 6 - > 2 C 3H 4O 3 + 2 ATP + 2 NADH + 2 H + NADH, o NAD + en su estado des-protonado (dinucleótido de nicotinamida y adenina), es un llamado portador de electrones de alta energía y un intermediario en muchas reacciones celulares involucradas en la liberación de energía. Tenga en cuenta dos cosas aquí: una es que la glucólisis sola no es tan eficiente en la liberación de ATP como lo es la respiración aeróbica completa, en la cual el piruvato producido en la glucólisis ingresa al ciclo de Krebs en el camino hacia los átomos de carbono que aterrizan en la cadena de transporte de electrones. Mientras que la glucólisis tiene lugar en el citoplasma, las reacciones posteriores de la respiración aeróbica se producen en orgánulos celulares llamados mitocondrias. Glucosa, que contiene una estructura de seis anillos que incluye cinco átomos de carbono y uno átomo de oxígeno, es transportado a la célula a través de la membrana plasmática por proteínas de transporte especializadas. Una vez dentro, se fosforila inmediatamente, es decir, se le une un grupo fosfato. Esto hace dos cosas: le da a la molécula una carga negativa, de hecho atrapándola dentro de la célula (las moléculas cargadas no pueden atravesar fácilmente la membrana plasmática) y desestabiliza la molécula, configurándola más realidad dividida en componentes más pequeños. br> La nueva molécula se llama glucosa-6-fosfato (G-6-P), ya que el grupo fosfato está unido al átomo de carbono número 6 de glucosa (el único que se encuentra fuera de la estructura del anillo). La enzima que cataliza esta reacción es una hexoquinasa; "hex-" es el prefijo griego para "seis" (como en "azúcar de seis carbonos") y las quinasas son enzimas que deslizan un grupo fosfato de una molécula y lo fijan en otro lugar; en este caso, el fosfato se toma del ATP, dejando ADP (difosfato de adenosina) a su paso. El siguiente paso es la conversión de glucosa-6-fosfato a fructosa-6-fosfato (F-6- PAG). Esto es simplemente una reorganización de átomos, o una isomerización, sin adiciones o sustracciones, de modo que uno de los átomos de carbono dentro del anillo de glucosa se mueve fuera del anillo, dejando un anillo de cinco átomos en su lugar. (Puede recordar que la fructosa es "azúcar de fruta", un elemento dietético común y natural). La enzima que cataliza esta reacción es la fosfoglucosa isomerasa. El tercer paso es otra fosforilación, catalizada por la fosfofructoquinasa (PFK) y produciendo fructosa 1,6-bisfosfato (F-1,6-BP). Aquí, el segundo grupo fosfato se une al átomo de carbono que se extrajo del anillo en el paso anterior. (Consejo de nomenclatura química: la razón por la que esta molécula se llama "bisfosfato" en lugar de "difosfato" es que los dos fosfatos están unidos a diferentes átomos de carbono, en lugar de que uno se una al otro frente a un enlace carbono-fosfato). así como el paso de fosforilación anterior, el fosfato suministrado proviene de una molécula de ATP, por lo que estos primeros pasos de la glucólisis requieren una inversión de dos ATP. El cuarto paso de la glucólisis rompe una molécula de seis carbonos ahora altamente inestable. en dos moléculas diferentes de tres carbonos: gliceraldehído 3-fosfato (GAP) y dihidroxiacetona fosfato (DHAP). La aldolasa es la enzima responsable de esta escisión. Puede discernir a partir de los nombres de estas moléculas de tres carbonos que cada una de ellas obtiene uno de los fosfatos de la molécula original. Con la glucosa manipulada y dividida en aproximadamente igual debido a un pequeño aporte de energía, las reacciones restantes de la glucólisis implican la recuperación de los fosfatos de una manera que resulta en una ganancia de energía neta. La razón básica de que esto ocurra es que eliminar grupos fosfato de estos compuestos es más energéticamente favorable que simplemente tomarlos directamente de las moléculas de ATP y aplicarlos a otros fines; piense en los pasos iniciales de la glucólisis en términos de un viejo adagio: "Hay que gastar dinero también para ganar dinero". Al igual que G-6-P y F-6-P, GAP y DHAP son isómeros: Tienen la misma fórmula molecular, pero diferentes estructuras físicas. Como sucede, GAP se encuentra en la vía química directa entre la glucosa y el piruvato, mientras que DHAP no. Por lo tanto, en el quinto paso de la glucólisis, una enzima llamada triose fosfato isomerasa (TIM) se hace cargo y convierte DHAP en GAP. Esta enzima se describe como una de las más eficientes en todo el metabolismo energético humano, acelerando la reacción que cataliza por un factor de aproximadamente diez mil millones (10 10). En el sexto paso, GAP se convierte en 1,3-bisfosfoglicerato (1,3-BPG) bajo la influencia de la enzima por la gliceraldehído 3-fosfato deshidrogenasa. Las enzimas deshidrogenasa hacen exactamente lo que sugieren sus nombres: eliminan átomos de hidrógeno (o protones, si lo prefiere). El hidrógeno liberado de GAP encuentra su camino hacia una molécula de NAD +, produciendo NADH. Tenga en cuenta que a partir de este paso, a efectos contables, todo se multiplica por dos, ya que la molécula inicial de glucosa se convierte en dos La reversión de facto de las reacciones de fosforilación anteriores de la glucólisis comienza con el séptimo paso. Aquí, la enzima fosfoglicerato quinasa elimina un fosfato de 1,3-BPG para producir 3-fosfoglicerato (3-PG), con el fosfato aterrizando en ADP para formar ATP. Dado que, nuevamente, esto involucra dos moléculas de 1,3-BOG por cada molécula de glucosa que ingresa a la glucólisis aguas arriba, esto significa que se producen dos ATP en general, cancelando los dos ATP invertidos en los pasos uno y tres. En paso ocho, 3-PG se convierte en 2-fosfoglicerato (2-PG) gracias a la fosfoglicerato mutasa, que extrae el grupo fosfato restante y lo mueve un carbono. Las enzimas mutasa difieren de las isomerasas en que, en lugar de reorganizar significativamente la estructura de una molécula completa, simplemente desplazan un "residuo" (en este caso, un grupo fosfato) a una nueva ubicación mientras dejan la estructura general intacta. En el paso nueve, sin embargo, esta preservación de la estructura se vuelve discutible, ya que 2-PG se convierte en piruvato de fosfoenol (PEP) por la enzima enolasa. Un enol es una combinación de un alk_ene_ y un alcohol. Los alquenos son hidrocarburos que incluyen un doble enlace carbono-carbono, mientras que los alcoholes son hidrocarburos con un grupo hidroxilo (-OH) adjunto. El -OH en el caso de un enol se une a uno de los carbonos involucrados en el doble enlace carbono-carbono de la PEP. Finalmente, en la décima y última etapa de la glucólisis, la PEP se convierte en piruvato por la enzima piruvato quinasa Si sospecha por los nombres de los diversos actores en este paso que se generan otras dos moléculas de ATP en el proceso (una por reacción real), está en lo correcto. El grupo fosfato se elimina de PEP y se agrega al ADP que acecha cerca, produciendo ATP y piruvato. El piruvato es una cetona, lo que significa que tiene un carbono no terminal (es decir, uno que no está al final de la molécula) involucrado en un doble enlace con oxígeno y dos enlaces simples con otros átomos de carbono. La fórmula química para el piruvato es C 3H 4O 3, pero expresar esto como (CH 3) CO (COOH) ofrece una imagen más esclarecedora del producto final de la glucólisis. La cantidad total de energía liberada (es tentador pero incorrecto decir "producido", ya que la "producción" de energía es un nombre inapropiado) se expresa convenientemente como dos ATP por molécula de glucosa. Pero para ser más matemáticamente precisos, esto también es 88 kilojulios por mol (kJ /mol) de glucosa, igual a aproximadamente 21 kilocalorías por mol (kcal /mol). Un lunar de una sustancia es la masa de esa sustancia que contiene el número de moléculas de Avogadro, o 6.02 × 10 23 moléculas. La masa molecular de la glucosa es de poco más de 180 gramos. Dado que, como se señaló anteriormente, la respiración aeróbica puede derivar más de 30 moléculas de ATP por glucosa invertida, es tentador considerar la producción de energía de la glucólisis sola como algo trivial , casi sin valor. Esto es completamente falso. Tenga en cuenta que las bacterias, que han existido por cerca de tres mil quinientos millones de años, pueden funcionar bastante bien usando la glucólisis sola, porque estas son formas de vida exquisitamente simples que tienen pocos de los requisitos que los organismos eucariotas tienen. De hecho, es posible ver la respiración aeróbica de manera diferente al poner todo el esquema sobre su cabeza: si bien este tipo de producción de energía es ciertamente una maravilla bioquímica y evolutiva, los organismos que la utilizan en su mayor parte dependen absolutamente de ella. Esto significa que cuando no se encuentra oxígeno en ninguna parte, los organismos que dependen exclusivamente o en gran medida del metabolismo aeróbico, es decir, todos los organismos que leen esta discusión, no pueden sobrevivir por mucho tiempo en ausencia de oxígeno. En cualquier caso, La mayoría del piruvato producido en la glucólisis se mueve hacia la matriz mitocondrial (análoga al citoplasma de células enteras) y entra en el ciclo de Krebs, también llamado ciclo del ácido cítrico o ciclo del ácido tricarboxílico. Esta serie de reacciones sirve principalmente para generar muchos portadores de electrones de alta energía, tanto NADH como un compuesto relacionado llamado FADH 2, pero también produce dos ATP por molécula de glucosa original. Estas moléculas luego migran a la membrana mitocondrial y participan en las reacciones en cadena de transporte de electrones que finalmente liberan 34 ATP más. En ausencia de oxígeno suficiente (como cuando hace ejercicio vigorosamente), parte del piruvato se somete fermentación, un tipo de metabolismo anaeróbico en el que el piruvato se convierte en ácido láctico, generando más NAD + para su uso en procesos metabólicos.
son comunes a prácticamente todas las células, tanto en organismos procariotas (casi todas las cuales son bacterias) como en organismos eucariotas (principalmente plantas, animales y hongos).
Glucólisis: reactivos y productos
Glucólisis: pasos iniciales
Glucólisis: pasos finales
moléculas de GAP. Así, después de este paso, dos moléculas de NAD + se han reducido a dos moléculas de NADH.
Consideraciones energéticas y el destino del piruvato