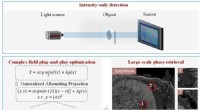
Motores moleculares caminando a lo largo de un microtúbulo. Crédito:M. Rank / PRL
Los polímeros filamentosos llamados microtúbulos juegan un papel vital en la segregación cromosómica y el transporte molecular. Un equipo de LMU ha examinado ahora cómo varían las longitudes de los microtúbulos en respuesta a los cambios en la disponibilidad de sus componentes proteicos.
Las células poseen un esqueleto interno, lo que les permite alterar su forma y migrar activamente. Este citoesqueleto se compone de varios sistemas de filamentos, de los cuales los microtúbulos son uno. Los microtúbulos cilíndricos tienen paredes formadas por 13 protofilamentos, cada una de las cuales consta de subunidades heterodiméricas que contienen dos proteínas tubulinas relacionadas. Los microtúbulos no solo confieren estabilidad mecánica a las células y dictan sus formas, también sirven como una red de transporte intracelular. Es más, los microtúbulos son los componentes principales del huso mitótico, que media la segregación ordenada de los conjuntos de cromosomas replicados en las dos células hijas durante la división celular. Todas estas funciones requieren una regulación dinámica de la longitud de los microtúbulos. Un grupo de físicos de LMU dirigido por el profesor Erwin Frey, en colaboración con el profesor Stefan Diez (Universidad Técnica de Dresde y el Instituto Max Planck de Biología Celular Molecular y Genética, Dresde), Ahora han desarrollado un modelo en el que las proteínas motoras responsables del transporte de carga a lo largo de los protofilamentos también sirven para regular la longitud de los microtúbulos. El modelo está descrito y validado experimentalmente en la revista. Cartas de revisión física .
En un trabajo anterior, El grupo de Frey había demostrado que la densidad de los motores moleculares unidos a los filamentos influye en si el microtúbulo crece o se contrae. y que su efecto depende de la longitud del filamento en cuestión. Cuanto más largo sea el microtúbulo, cuanto mayor sea el número de proteínas motoras que puede acomodar. Las moléculas motoras llamadas cinesinas avanzan a lo largo del protofilamento, pasando de un dímero al siguiente. Cuando una proteína kinesina llega al final, se desprende del filamento llevando la tubulina a la que está unido. Como consecuencia, si la densidad del motor en el protofilamento es alta, la contracción continuará. Por otra parte, un nuevo dímero de tubulina puede unirse al final. Al final, Por tanto, la contracción dependiente del motor compite con el crecimiento de los microtúbulos. "Por eso, asumiendo que los recursos (es decir, tanto tubulinas como motores moleculares) están presentes en el acceso, Habrá una longitud de filamento a la que se equilibran las tasas de crecimiento y contracción, "dice Matthias Rank, primer autor del estudio. Sin embargo, en una celda real, Es poco probable que estos componentes estén disponibles en cantidades ilimitadas. Por ejemplo, La formación del huso mitótico agota significativamente el número de moléculas de tubulina libres en la fase soluble del citoplasma. En el nuevo estudio, los investigadores exploraron los efectos de tal limitación de recursos en la regulación de la longitud de los microtúbulos.
Usando simulaciones basadas en un modelo matemático de dinámica de polímeros, encontraron que bajo estas condiciones entran en juego dos mecanismos distintos de regulación de la longitud. Cuál de estos se vuelve dominante depende de las concentraciones relativas de las tubulinas y las proteínas motoras:en un cierto rango de concentración, el equilibrio dinámico entre el crecimiento y la contracción de los microtúbulos funciona como lo haría si los recursos no fueran limitantes. "Pero las cosas son diferentes cuando uno de los recursos necesarios escasea", dice Rank. "Ese es el caso, por ejemplo, cuando no hay suficientes moléculas motoras disponibles para desencadenar una rápida despolimerización de los protofilamentos ". En esta situación, los microtúbulos continúan creciendo hasta que la concentración de tubulinas cae por debajo de un valor crítico. Es más, hay un rango de concentración en el que ambos procesos están activos. "En este caso, observamos que los microtúbulos vienen en dos tamaños y que a veces cambian entre las dos longitudes ", dice Frey. "En términos físicos, esto puede describirse como una transición de fase ". Los experimentos in vitro llevados a cabo por su coautor en Dresde han confirmado la existencia de este régimen de transición predicho por el modelo de Munich. El equipo está convencido de que sus resultados también son aplicables a otros sistemas de polímeros , y sospechan que la limitación de recursos clave puede desempeñar un papel importante en la regulación de otros procesos celulares.